Creationtriumphs_05
CHAPTER FIVE
Darwinian Evolution —
Fact or Theory?
The exponents of Darwin's evolution have long declared this theory a scientific fact. But by its own rules, science requires empirical proof—that is, observation. After over 140 years of research, what empirical proof have Darwinists compiled? The following discussion applies the litmus test of science itself against the various speculations evolutionists have pursued for explaining the evolutionary process.
Natural Selection Vs. Artificial Selection
"Natural selection" proposes species are constantly replacing species in a process called "descent with modification." Natural selection is the mechanism responsible for all the varieties of plants and animals. The guiding force—"survival of the fittest"— is blindly deciding which species survive.
What Darwin identified as "variation" is today explained as achieved by mutations. "Mutations are randomly occurring changes which are nearly always harmful when they produce effects in the organism large enough to be visible, but which may occasionally slightly improve the organism's ability to survive and reproduce." (1)
But did Darwin explain his theory of evolution by natural selection? The noted Darwinist, Douglas Futuyma, explains:
When Darwin wrote THE ORIGIN OF SPECIES, he could offer no good cases of natural selection because no one had looked for them. He drew instead an analogy with the artificial selection that animal and plant breeders use to improve domesticated varieties of animals and plants. By breeding only from the woolliest sheep, the most fertile chickens, and so on, breeders have been spectacularly successful in altering almost every imaginable characteristic of our domesticated animals and plants to the point where most of them differ from their wild ancestors far more than related species differ from them. (2)
Do all evolutionists cite artificial selection as a proof of what "natural selection" is supposed to achieve? The eminent French zoologist Pierre Grasse, an evolutionist but a strong anti-Darwinist, concluded that the results of artificial selection pro-vide powerful testimony against Darwin's theory:
In spite of the intense pressure generated by artificial selection (eliminating any parent not answering the criteria of choice) over whole millennia, no new species are born. A comparative study of sera, hemoglobins, blood proteins, interfertility, etc., proves that the strains remain within the same specific definition. This is not a matter of opinion or subjective classification, but a measurable reality. The fact is that selection gives tangible form to and gathers together all the varieties a genome is capable of producing, but does not constitute an innovative evolutionary process. (3)
In other words, the reason that dogs do not become as big as elephants, much less change into elephants, is not that we have not been breeding them long enough. Dogs do not have the genetic capacity for that degree of change, and they stop getting bigger when the genetic limit is reached. Cries of "not enough time" to produce new species should be muted by exhaustive research with the fruitfly. Since the life span of the fruitfly is so short, it represents mutation observation over thousands of generations in a short period of time. The fact that scientists have been able to breed fruitflies into every possible genotype only proves that fruitflies can be caused to change through artificial selection, but not natural selection. If artificial selection proves anything, it proves that an intelligent manipulation of genetics is sometimes able to produce a woollier sheep, a better tomato, and a different looking fruitfly. In any case, the end result of all these genetic experiments is that a fruitfly is still a fruitfly—not a new species. This does not make a case for beneficial mutations being the engine behind natural selection.
Tautology is a way of saying the same thing twice. The noted law professor turned prosecutor of Darwinism, Phillip E. Johnson, zeroed in on the tautology fallacy of Darwinism as reflected in the following:
The famous philosopher of science Karl Popper at one time wrote that Darwinism is not really a scientific theory because natural selection is an all-purpose explanation which can account for anything, and which, therefore, explains nothing. Popper backed away from this position after he was besieged by indignant Darwinist protests, but he had plenty of justification for taking it. As he wrote in his own defense, "some of the greatest contemporary Darwinists themselves formulate the theory in such a way that it amounts to the tautology that those organisms that leave most offspring leave most offspring," citing Fisher, Haldane, Simpson, "and others." (4)
One of the "others" referred to was Waddington, whose explanation Johnson said should be preserved for posterity:
Darwin's major contribution was, of course, the suggestion that evolution can be explained by the natural selection of random variations. Natural selection, which was at first considered as though it were a hypothesis that was in need of experimental or observational confirmation, turns out on closer inspection to be a tautology, a statement of an inevitable but previously unrecognized relation. It states that the fittest individuals in a population (defined as those which leave most offspring) will leave the most offspring. This fact in no way reduces the magnitude of Darwin's achievement; only after it was clearly formulated, could biologists realize the enormous power of the principle as a weapon of explanation. (5)
Johnson observed that this statement of Waddington's was not just an offhand statement:
That was not an offhand statement, but a considered judgment published in a paper presented at the great convocation at the University of Chicago in 1959 celebrating the hundredth anniversary of the publication of THE ORIGIN OF SPECIES. Apparently, none of the distinguished authorities present told Waddington that a tautology does not explain anything. When I want to know how a fish can become a man, I am not enlight-ened by being told that the organisms that leave the most off-spring are the ones that leave the most offspring. The important point is that the Darwinists have been tempted continually by the thought that their theory could be given the status of an a priori truth, or a logical inevitability; so that it could be known to be true without the need of empirical confir-mation. Their susceptibility to this temptation is understandable. When the theory is stated as a hypothesis requiring empirical confirmation, the supporting evidence is not impressive. (6)
Thus, many of the scientific community hold natural selection as a philosophical necessity—some scientists demand a natura-listic explanation for everything. Since God or any other "vital force" that drives evolution is excluded by the National Academy of Sciences, evolutionists have to make do with what is left when the unacceptable has been excluded. Natural selection is the best of the remaining alternatives—probably the only alternative.
There are many other anomalies inherent in the idea of natural selection. Why haven't many of the "lower-order" creatures with us today evolved into something on the same level as humans? Why do some animals risk their own safety to warn others of an approaching predator? Why is a female pea-hen attract-ed to a male peacock with "life-threatening" decorations? Explanations for these inconsistencies make it difficult to conceive of a way to test the claims empirically.
One of the concepts related to mutations that evolutionists try to avoid is the evidence of "saltations," that is, sudden leaps by which a new type of organism appears in a single generation. Darwin himself thought that saltations were nothing less than miracles. T. H. Huxley warned Darwin of dismissing saltation too quickly. Huxley's reason for this caution was the lack of fossil record supporting the gradualism Darwin proposed. Problems are created when evolutionists discard the concept of saltation. Why would wings or eyes continue to develop in a creature with no apparent functionality—unless the system evolved at once? The first step towards a new function—such as vision or ability to fly—would not necessarily provide any advantage unless the other parts required for the function appeared simultaneously.
A noted professor, Richard Goldschmidt, challenged the Darwinian concept of micro-mutations, holding that this concept "could account for no more than variations within the species' boundary." (7) He admitted that macro-mutation would usually produce harmful development, but thought it possible that occasionally a "hopeful monster" would emerge and develop a new species. But with what mate is the logical question.
The micro-macro question has been argued by two contemporary evolutionist giants, Dawkins and Gould. Richard Dawkins, the Dean of British Scientists, defended Darwin's gradualism— micro-mutation. Stephen Jay Gould, the eminent paleontologist of Harvard, attempted to harmonize saltations with a form of macro-mutation.
The bottom line is that there is no way to prove if either such mutations ever occurred. If after "massive" research, scientists were able to alter the genetic code of a fish to produce an amphibian, would that prove anything? No. This artificial manipulation proves nothing about random changes. This type of experimentation would only prove that these changes could be planned and executed by an intelligent scientist. But whether God as the Intelligent Creator would employ this method to achieve wonderful variety is another question.
Demonstrating that mutations can be beneficial poses a significant problem to the evolutionist. The mathematical calculations required to predict whether micro- or macro-mutations would be advantageous are staggering. Mathematician D. S. Ulam concluded that the amount of mutations needed to create an eye made it impossible. Evolutionists retorted by stating that the eye had evolved. Ernst Mayr responded, "Somehow or other by adjusting these [Ulam's] figures, we will come out all right. We are comforted by the fact that evolution has occurred." (8) This attitude in the scientific community of evolutionists reflects an incredible position. Notwithstanding the complete lack of empirical evidence, evolution is considered an a priori fact and law.
The lack of fossil evidence is perhaps the greatest challenge to Darwinism. Darwin himself bemoaned the fact that we did not "everywhere see innumerable transitional forms." He even admitted that the state of fossil evidence was "the most obvious and gravest objection which can be urged against my theory." (9)
After 140 years of evolutionists desperately looking for missing links, Gould offered "punctuated equilibrium" to deal with the embarrassing fact: "The fossil record today on the whole looks very much as it did in 1859." (10) Gould summarized his concept as follows:
The history of most fossil species includes two features particularly inconsistent with gradualism:
1. Stasis. Most species exhibit no directional change during their tenure on earth. They appear in the fossil record looking pretty much the same as they disappear; morphological change is usually limited and directionless.
2. Sudden appearance. In any local area, a species does not arise gradually by the steady transformation of its ancestors; it appears all at once and "fully formed." (11)
The essential point of Gould's "punctuated equilibrium" is that the new species appear in peripheral groups, whereas the fossil record is available basically from the larger, main populations— that is why new species seem to appear suddenly:
Speciation (the formation of new species) occurs rapidly, and in small groups which are isolated on the periphery of the geo-graphical area occupied by the ancestral species.... Because fossils are mostly derived from large, central populations, a new species would appear suddenly in the fossil record following its migration into the center of the ancestral range. (12)
In this small isolated population, Gould explained, selective pressures might cause favorable variations to spread more rapidly. In this manner, a new species would arise in the peripheral
area without leaving fossil evidence. "Punctuated equilibrium" is a very interesting speculation. But how much more plausible to explain by the Genesis account what is already empirically self-evident from the fossil record! Well defined species were created in a logical sequence of complexity, each allowing for genetic variations within its own "kind."
The single greatest challenge which the fossil record poses for Darwinism is the "Cambrian Explosion" which they date around 600 million years ago. Nearly all the animal phyla appear in the rocks of this Cambrian period without a trace of the evolutionary ancestors that Darwinism requires. As Richard Dawkins put it, "It is as though they were just planted there, without evolutionary history." In fact, Darwin himself found no evidence of the existence of pre-Cambrian life and conceded in THE ORIGIN OF SPECIES, "The case at present must remain inexplicable, and may be truly urged as a valid argument against the view here entertained." If his theory were true, Darwin wrote, the pre-Cambrian world must have "swarmed with living creatures." But, as Dawkins observed, after over a hundred years of searching the fossil record, the pre-Cambrian world did not swarm with living creatures.
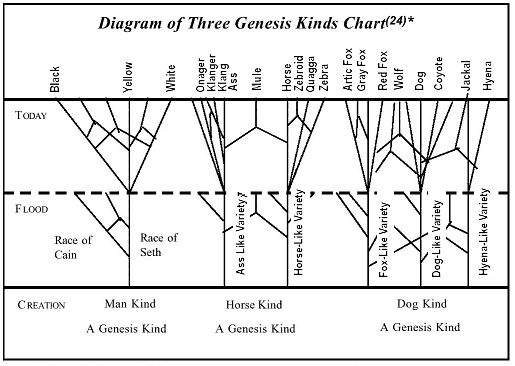
Missing links haunt evolutionists throughout the fossil record. Though there is evidence of micro-mutation within the Genesis kind, after 140 years of gene manipulation in the laboratory and intensive investigation of the fossil record, there is no evidence of micro- or macro-mutation bridging from one Genesis kind to another. Darwinians and neo-Darwinians are frantically advancing multiple theories to minimize this lack of empirical fossil evidence. Although they present a united front that evolution is a fact, they are in vast disarray when it comes to the how of proving that evolution is even a workable possibility. In the face-off between the two Darwinian giants, Dawkins and Gould, each claimed that his own mutation theory—voiding out any other—is the only way to explain missing links. The truth is Dawkins' and Gould's evaluations of each other's mutation theories cancel out each other! Neither view can provide empirical evidence of bridging gaps in the fossil record. But the sudden appearances of new species in the fossil record—the "Cambrian Explosion"—is consistent with the Creation model. The fossil record is still testimony against Darwinian evolution.
Commenting on the fossilized skeleton of Leonardo, a recently discovered, mummified, brachylophosaurus dinosaur, Nate Murphy, curator of paleontology at the Phillips County Museum, Montana., conceded:
"Paleontology is not an exact science. All we have are bones, and from there we develop theories about what the animals looked like, how they moved, and what they ate. A specimen like Leonardo will take a lot of guess work out and really tell us if Steven Spielberg's getting it right." (13)
Some evolutionists attempt to establish their theory by pointing out certain similarities embryos share that belong to different species. Since evolution is considered a fact, biological relationships are assumed to signify evolutionary relationships. Homology and embryology have been put forward as proof of the "fact of evolution."
In 1866 Ernst Haeckel formulated what came to be know as the "Biogenetic Law" or "Haeckel's Law," which simply stated means, an embryo will recapitulate (summarize) the evolutionary stages of a life form during the embryo's development. To Darwin, Haeckel's Law established the fact of evolution. Therefore, Darwin based his research in the fields of natural selection, the fossil record, the vertebrate sequence, pre-biological evolution, etc., on the a priori assumption that evolution was a fact because of this "Law." This a priori "logic" has been the basic flaw of evolutionists ever since.
For centuries philosophers have noted the relationships between different animals and always attributed these similarities—not as inheritance from common ancestors—but to a sort of blueprint called the "Archetype," which existed only in some metaphysical realm, such as the mind of the Divine Creator. Darwin's theory of "descent with modification" offered a naturalistic alternative to the idea of the Archetype.
Darwin described his theory, "The characters which naturalists consider as showing true affinity between any two or more species, are those which have inherited from a common parent, and insofar, all true classification is genealogical." (14)
Neither laboratory science nor the fossil record has been able to provide empirical evidence for the theory of "descent with modification." However, the "fact of evolution" seems to stand unscathed regardless of the lack in the validity of its records and proposed mechanisms. Stephen Jay Gould made a false analogy regarding the "fact of evolution":
Facts are the world's data. Theories are structures of ideas that explain and interpret facts. Facts do not go away while scientists debate rival theories for explaining them. Einstein's theory of gravitation replaced Newton's, but apples did not suspend them-selves in mid-air pending the outcome. And human beings evolved from ape-like ancestors whether they did so by Darwin's proposed mechanism or by some other, yet to be identified. (15)
Is this comparison fair? Johnson ably refuted Gould's reasoning:
The analogy is spurious. We observe directly that apples fall when dropped, but we do not observe a common ancestor for modern apes and humans. What we do observe is that apes and humans are physically and biochemically more like each other than they are like rabbits, snakes, or trees. The ape-like common ancestor is a hypothesis in a theory, which purports to explain how these greater and lesser similarities came about. The theory is plausible, especially to a philosophical materialist, but it may nonetheless be false. (16)
Without empirical evidence in the laboratory or fossil records, Darwin and his loyal legion seized upon homology in embryology to prove the "fact" of evolution. The argument from embryology is based primarily upon the ideas of Haeckel's biogenetic law. In erudite descriptive form, this law means "ontogeny recapitulates phylogeny." Or, in other words, an "individual" will summarize his evolutionary history by passing through similar evolutionary stages during his embryological development.
So it has been popularly believed that man has a gill stage, a hair stage, tail stage, protozoan stage, worm stage, etc. Embryo similarities are an evidence all are taught to believe even in elementary biology courses. Surprising as it may seem, however, this evidence has been rejected by practically all competent biologists.
Notwithstanding his "fact-of-evolution" posture, Gould had to disassociate himself from Haeckel's Law—the only law of science that seemed to give credence that evolution was, indeed, a fact. Gould remarked:
. . .the New York public schools taught him Haeckel's doctrine, that ontogeny recapitulates phylogeny, fifty years ago it had been abandoned by science. . .behind closed doors many scien-tists will admit to thinking that "there really is something to it after all." (17)
Haeckel's Law has been replaced by Von Baer's Law which "asserts that resemblances among embryos reflect levels of bio-logical classification, so that all vertebrates, for example, look very similar in early development but become increasingly dissimilar as they approach their adult forms." Unfortunately, the embryos do not start out similarly, but only converge to a similar embryo in the middle of development only to diverge again to develop into fish, birds and mammals.
Thus, while all vertebrates pass through an embryonic stage in which there is a resemblance, actually they develop to this stage very differently. When the egg is fertilized, each Genesis kind follows its own plan of embryonic development. Fish, amphibians,birds and mammals each follows their own pattern. Only by ignoring these early stages of development can Darwin's theory harmonize with the facts of embryology. Yet it was the early stages that were crucial to Darwin's claim. The latter stages of embryo development are also out of sync with Darwinian expectations. For instance, limb development is an instructive example. The embryonic development of limb bones reveals patterns of division, branching and cartilage production which differ from Genesis kind to Genesis kind without conforming to predictions based on the theory of common descent.
The "fact of embryology" is that all vertebrate embryos follow different patterns of development, then midway through the process converge into similar appearances—and again diverge until they finally through diverse processes develop similar bone structure in their limbs. Can embryology be harmonized with either a Creator's "archetype" or Darwin's "descent with modification"? That embryology alone cannot be used to prove either is fact. However, the scale tips more in support of creation, as Johnson observed:
If embryology is our best guide to genealogy, as Darwin thought, our guide seems to be telling us that vertebrates have multiple origins and did not inherit their similarities from a common ancestor. (18)
Imposing vertebrate exhibits in museums show neat progress sive evolutionary sequences of vertebrate development from the simple to the complex. The evolutionist smugly says, "There you have it—proof positive that evolution is a fact!" Marsh's fossil pedigree of the horse displayed at Yale University convinced Thomas H. Huxley himself of the "irrefutable truth" of evolution. There it was—the evolution of the horse beginning with Eohippus (the so-called "Dawn Horse") which was the size of a fox terrier, possessing several toes with the display climaxing with Equus, the tall, majestic modern-day horse. Darwin had planned to make the trip to see Marsh's collection, but health did not permit. As P. I. Lull lamented, "He died without having seen such a culminating proof of his theory of evolution." But there is less than meets the eye on these impressively neat simple-to-complex, small-to-large displays of vertebrate sequences! This "less than meets the eye" was quite evident in an interview with Gareth Nelson, of the American Museum of Natural History. When asked about the question of vertebrate sequence, Nelson said, "We've got to have some ancestors. We'll pick those. Why? Because we know they have to be there, and these are the best candidates. That's by and large the way it has worked. I am not exaggerating." (19)
Again, this is not empirical proof. It is the same persistent flaw of the evolutionist—the a priori assumption. If evolution is assumed as fact, then the vertebrate sequence has to be rigged to prove evolution. The sequence from small several-toed ancestors to large one-toed horses is nowhere to be found in the fossil record. Furthermore, many contradictions to this presumed order are conspicuous. For example, two modern-type horses, Equus nevadensis and Equus occidentalis, have been found in the same geological stratum as Eohippus. This fossil record verifies modern- day-type horses were grazing side by side with their so-called ancestor.
An evolutionist of note, G. G. Simpson, asserted that the development of the horse is not by "orthogenesis"—in a straight line. Simpson's vertebrate sequence of the horse is vastly different from Marsh's at Yale. (Simpson was from Harvard.) Simpson declared, "This is not a sequence involving lower and higher zones, but evolution in a single, changing zone." (20) Regarding the gradual reduction from several toes to a single toe or hoof, Simpson said that it is "flatly fictitious." (21)
Horses vary today from "Gumbo," an 18-inch tall American Miniature Yearling Stallion (which is even smaller in Argentina)—to the seven-foot high 3,200-pound Clydesdales. Another contradiction to the presumed order of evolving horses is that some present-day Shire horses have more than one toe per foot. (22) Similarly, antelopes vary from the 12-inch Suni to the 6- foot, 2,000-pound Eland.
Evolutionists take great satisfaction in discussing evolution within the horse specie. But what type of evolution is this? "Micro-evolution" at best—wonderful variation within a Genesis kind—the horse "kind." The evolutionists' forced extrapolation that the great diversity within the horse kind proves Darwinism evolution is not valid. By definition, Darwinism requires evolu-tion from one Genesis kind to another Genesis kind.
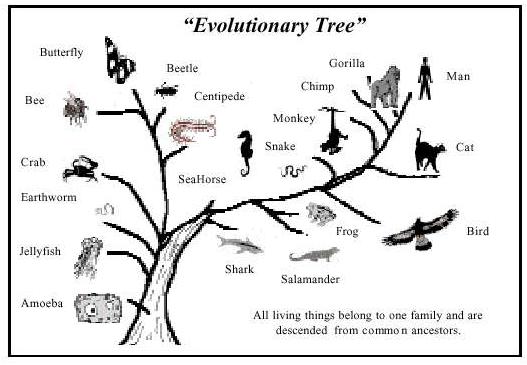
How can documentation of evolution of horses into horses, moths into moths, minulus into minulus, fruitflies into fruitflies, prove the evolution from amoeba to man? At every stage of the evolutionary tree, there are question marks demanding proof for common ancestry of all organism. (See the diagram on the next page.)
Fish to Amphibians. Some Darwinists propose the rhipidistians, an extinct order of fish, as a possible "ancestral group." The rhipidistians are thought to have skeletal features similar to early amphibians which have bones that look like they have the potential to evolve into legs. Not so! said evolutionist Barbara J. Stahl in her comprehensive textbook:
None of the known fish [sic] is thought to be directly ancestral to the earliest land vertebrates. Most of them lived after the first amphibians appeared, and those that came before show no evidence of developing the stout limbs and ribs that characterized the primitive tetrapods. (23)
The coelacanth, an ancient fish thought to be extinct in the same class as rhipidistians, was caught in the Indian Ocean. When dissected, its skeletal features and internal organs showed no signs of being preadapted for a land environment. There is still no evidence of any transition form of life between the fish and the amphibian classifications.
Amphibians to Reptiles. Transitional ancestors to the reptiles were required. Darwinists selected the so-called stem reptile, Seymouria. Embarrassment was readily imagined when this selected "ancestor" was dated by evolutionary methods as having existed 20 million years after reptiles already appeared on the earth. Evolutionists do not present a valid case for any possible link between amphibians and reptiles.
Reptiles to Mammals. If reptiles, indeed, evolved into mammals, transitional ancestors for mammals would need to be established. Evolutionists chose the large order of therapsida, a mammal- like reptile. As a recognized expert in mammal-like reptiles, . Hopson ventured a vertebrate sequence of therapsids to bridge from different orders and subgroups of reptiles ending his equence with a mammal—the Morgamicodon. The only problem as that the Morgamicodon was substantially older than the therapsid that preceded it! (25) This attempt hardly qualifies as an ancestry hypothesis.
In any case, more than one transitional life form would be necessary to establish transitional ancestry because of so much diversity among mammals. As Johnson observes:
The mammal class includes such diverse groups as whales, porpoises, seals, polar bears, bats, cattle, monkeys, cats, pigs, and opossums. If mammals are a monophyletic group, then the Darwinian model requires that every one of the groups have descended from a single unidentified small land mammal. Huge numbers of intermediate species in the direct line of transition would have had to exist [for every diverse group of mammals], but the fossil record fails to record them. (26)
Reptiles to Birds. In 1998 two fossils of feathered dinosaurs were discovered in China's Liaoning province. The fossils were acclaimed as the "missing link" between reptiles and birds. However, the feathers found on the two species, Caudipteryx and Protoarchaeopteryx, were fully formed—a true "missing link" would reveal stages of development between scales and feathers. The dinosaur fossils are reportedly 120-145 million years old, however, the oldest known bird fossil is the allegedly 150-mil-lion- year-old Archaeopteryx, a dead end side branch of the ancient avian line of birds. (27) Thus, the fossil sequence contradicts the conclusion that the bird evolved from the dinosaur—the bird fossils are older than the dinosaur fossils.
Apes to Humans. An anthropologist who believes in the evolution of humans from apes would select ancestors that would fit a neat sequence, even if these sequences are only constructed from a tooth or jawbone. The late Solly Zuckerman (now Lord Zuckerman), one of Britain's most influential scientists and leading primate experts, was an ardent evolutionist. Questioning the reliability of anthropology, he said that anthropology "is so astonishing that it is legitimate to ask whether much science is yet found in this field at all." (28) The evolutionary sequence attempts between apes and humans, Zuckerman admitted, "depend. . .partly on guesswork, and partly on some preconceived conception of the course of hominid evolution." (29)
If it is assumed in advance that ancestors of humans must have existed, there are only a few ambiguous examples of possible candidates for the transitional forms. These inconclusive examples represent what 140 years of frenzied research have produced. These sequences in the "evolutionary tree" are but scrawny branches when reason would demand numerous, even thick bushy transitional branches.
In addition, the resolute claims of Darwinian evolution beg the question, Why does not the fossil record abound with numerous species possessing partially formed organs, such as, 20 per-cent feather, 80 percent scale, 75 percent wing, 25 percent leg, 60 percent foot, 40 percent fin, 12 percent flower or 88 percent spore?
If evolution were a fact, then life today should still abound with these transitional organisms. Because evolutionists attempt to ignore this lack of empirical evidence in the fossil record, evolution remains just a theory without observable proof.
How did evolution allegedly begin in the first place? In a rather tentative letter, Charles Darwin in 1871 first proposed pre-biological evolution as follows:
It is often said that all the conditions for the first production of a living organism are now present, which could ever have been present. But if (and oh! what a big if!) we could conceive in some warm little pond, with all sorts of ammonia and phosphoric salts, lights, heat, electricity, etc., present, that a protein com-pound was chemically formed ready to undergo still more complex changes, at the present day such matter would be instantly devoured or absorbed, which would not have been the case before living creatures were formed. (30)
Robert Shapiro observed in 1986 that Darwin's offhand speculation "is remarkably current today, which is a tribute either to his foresight or our lack of progress." (31) A name for the theoret-ical model proposed by Alexander Oparin and J. B. S. Haldane in the 1920s caught journalistic imagination. The Oparin-Haldane Model became known as "prebiotic soup." Ever since, it has become an element of scientific folklore presented to the public in books and museum exhibits as the known source of life. But, as Johnson observes, "There is no reason to believe that life has a tendency to emerge when the right chemicals are sloshing about in a soup." (32)
The probabilities for life spontaneously exploding onto the scene are extremely small. The total probability of forming the proteins and DNA necessary and then transforming them into the first living entity—given astronomically large quantities of reagents and time—is 1/10 167,626 . (33) Without an intelligent Creator, life's probability is zero.
Fred Hoyle, considered by many the dean of cosmology as well as former long-time atheist, makes a good analogy that brings the problem to understandable terms. The chances of life
coming from prebiotic soup, he says, have the same probability of occurring that a "tornado sweeping through a junkyard might assemble a Boeing 747 from the materials therein." (34)
After the highly overrated Mill-Urey "prebiotic soup" experiment in the 1950s, various diverse models of prebiological evolution have been attempted or theorized. Even computer design models, called "spontaneous self-organization," have attempted to mimic the origin of life and its subsequent evolution. What have been the results of this tenacious research? The biological scientific community is not convinced. The respected periodical, Science, evaluated the computer models as follows:
Advocates of spontaneous organization are quick to admit that they aren't basing their advocacy on empirical data and laboratory experiments, but on abstract mathematics and novel computer models. The biochemist G. F. Joyce commented: "They have a long way to go to persuade mainstream biologists of the relevance [of this work]." (35)
Gerald F. Joyce observed in Nature that origin-of-life researchers have grown accustomed to a "lack of relevant experimental data." (36) A chemist with stature in the field, Robert Shapiro, candidly revealed that "the problems of explaining the origin of life have often been underestimated as investigators have exaggerated the importance of minor successes. . .[He affirmed] the existence of a naturalistic solution as a matter of faith." Robert Shapiro commented, "We have reached a situation where a theory has been accepted as a fact by some, and possible contrary evidence is shunted aside. This condition, of course, again describes mythology rather than science." (37) A leading figure in prebiological evolution, Director of the Institute for Biochemistry at the Johannes Gutenberg University in Mainz, Germany, Klause Dose commented: "At present all discussions on principal theories and experiments in the field either end in stalemate or in a confession of ignorance." (38)
Darwinists admit that to date there has been no evidence to validate any geochemical remnants of prebiotic molecules. This admission undermines the whole theory of evolution. Instead, the evidence is that all the carbonaceous deposits recovered from the oldest rocks are, without exception, the by-product of biological activity (as opposed to chemical evolution). Fully consistent with the discovery of life's by-products is the discovery of fossilized bacteria, cyanobacteria, about 3.5 billion years old, found in the oldest rocks yet discovered on Earth, dating around 3.9 billion years. (39,40)
Peter Ward, Professor of Geological Sciences at the University of Washington in Seattle, and Donald Brownlee, Professor of Astronomy at the University of Washington in Seattle and leader of the NASA Stardust mission, summarized:
...as we learn more about the nature of our planet's early environments, tranquil ponds or tide pools seem less and less likely to be plausible sites for the first life, or even to have existed at all on the surface of the early Earth. What Darwin could not appreciate in his time (nor could Haldane and Oparin, for that matter) was that the mechanisms leading to the accretion of Earth (and of other terrestrial planets) produced a world that, early in its history, was harsh and poisonous, a place very far removed from the idyllic tide pool or pond envisioned in the nineteenth and early twentieth centuries. In fact, we now have a very different view of the nature of the early Earth's atmosphere and chemistry. It is widely believed among planetary scientists that carbon dioxide, not ammonia and methane, dominated the earliest atmosphere and that the overall conditions may not have favored the widespread synthesis of organic molecules on Earth's surface. (41)
Norman Pace, one of the great pioneering microbiologists, admitted:
It seems fairly clear now that the early earth was, in essence, a molten ball with an atmosphere of high-pressure steam, carbon dioxide, nitrogen, and other products of volcanic emissions from the differentiating planet. It seems unlikely that any landmass would have reached above the waves (of a global ocean) to form the "tide pools" invoked by some theories for the origin of life. (42)
The presence of microbes thriving in 400 o C. hydrothermal ocean floor vents suggests to some biologists where life began. Laboratory experiments indicate prebiotic molecules can form under deep-sea vent conditions. (43) As researchers concede Earth's early atmosphere could not support prebiotic molecule formation, the appeal to deep-sea vents as a source of prebiotic molecules becomes even more important. (44)
However, for life to originate in this environment, ammonia must be present. Laboratory experiments at Penn State and SUNY-Stony Brook recently demonstrated the unlikelihood of ammonia formation under primitive hydrothermal vent conditions. Ammonia production occurs far too slowly in insufficient quantities to sustain prebiotic molecule formation. (45)
Inadequate ammonia production eliminates another possible source of prebiotic molecules, making the origin-of-life problem more intractable for naturalists. Without a source of prebiotic molecules, naturalistic origin-of-life pathways are blocked by additional barriers.
Contrary to the evolutionary theory that life in its minimal form is simple, evidence indicates to the contrary: life in its minimal form is chemically complex. Theoretical and experimental
work with the smallest known genome [the complete set of chromosomes necessary for reproduction], M. genitalium, indicates that life requires at least 250-350 gene products. (46-49)
Biophysicist Hubert Yockey has calculated the probability of forming a single gene product as one chance in 10 75 . Given this probability, Yockey calculated that if the hypothetical primordial soup contained about 10 44 amino acids, a hundred billion trillion years would yield a 95 percent chance for random formation of a functional protein only 110 amino acids in length (a single gene product). (50) If we assume that the universe is about 15 billion years old, less than one trillionth of the time has passed that would be needed to make even one of the 250-350 gene products necessary for minimal life, or one of the 1,500 gene products necessary for independent life.
Origin-of-life researchers must account not only for the simultaneous appearance of 250-350 gene products, but additionally for the remarkable internal organizational structure of bacteria at the protein level, both spatially and temporally. (51,52)
Not only does Darwinian evolution remain an unproven theory, but its advocates offer an incomplete, no-start theory. If there was no prebiological evolution to generate life in its simplest form, then life could not have evolved into ever-increasing com-plexity until it reached the current stature in man.
Scientific fact is only verifiable by the "scientific method," which by definition means, "the systematic pursuit of knowledge. . .through observation and experiment, and the formulation and testing of hypotheses." (53) The theory of evolution is disqualified by science's own round rules. What are the facts?
1. Evolutionists continually use micro-evolution, changes with-in a Genesis kind, to prove evolution. But this approach is not the point at issue. Darwinism requires macro-evolution from one Genesis kind to another Genesis kind. This "slight-of-hand" reasoning does not constitute empirical proof.
2. Darwinists invariably employ tautological reasoning or assume an a priori posture in claiming evolution to be factual. Neither type of reasoning provides empirical confirmation—and furthermore excludes the necessity of empirical testing. If macro-evolution is assumed to be a fact, rigging the fossil records, embryo misinterpretation or arbitrary vertebrate sequencing are the inevitable consequences.
3. Although Darwinists put up a united front to the public that evolution is a fact, some of the most damaging statements to this theory are advanced by Darwinists themselves. As they vie among themselves over personal theories, discrediting one another, they leave very little work for what?
One hundred forty years of intensive research to verify evolution has been to no avail. So why do Darwinists still tenaciously cling to this theory? The British evolutionist, D.M.S. Watson, unwittingly provided the answer:
The theory of evolution. . .is a theory universally accepted not because it can be proved by logically coherent evidence to be true, but because the only alternative, special creation, is clearlyincredible. (54)
As in Israel of old, those who cut down a tree, built an idol and then worshiped it, Darwinism prepared just such a hand-crafted idol (Isaiah 2:8; 44:13-17; 46:5-7). At its altar 99 percent of America's practicing scientists pay homage. Many are not Darwinist believers, but they dare not publicly profess otherwise, or they could be purged and shunned by America's top universities. In the sacred temples of Darwinism, academic freedom is a farce. For example, the veteran writer Forrest M. Mimms was dismissed by the noted periodical Scientific American simply because he did not believe in Darwin's evolution. . .never mind that he never mentioned this fact in his writings.
Great publishing houses like MacMillan, Doubleday and McGraw-Hill, do not dare publish anti-evolutionary works lest they rouse the ire of the scientific establishment. After all, they publish tens of thousands of scientific books annually for secondary and college level schools.
Self-deluded scientists cling desperately to the evolution theory, not because it is observable or verifiable, not because it is scientific, not because it is reasonable—but because they refuse to accept the only alternative, creation by God.
The Apostle Paul's words (Romans 1:20-22) reverberate down the centuries of time to our enlightened century:
For since the creation of the world His invisible attributes, His eternal power and divine nature, have been clearly seen, being understood through what has been made, so that they are with-out excuse. For even though they knew God, they did not honor Him as God, or give thanks, but they became futile in their speculations, and their foolish heart was darkened. Professing to be wise, they became fools... (NAS)